JEAN-PIERRE NADAL
Laboratoire de Physique Statistique de l'E.N.S.
Laboratoire associé au C.N.R.S. (U.R.A. 1306), à l'ENS, et aux Universités Paris VI et Paris VII
Ecole Normale Supérieure
24, rue Lhomond, F-75231 Paris Cedex 05, France
E-mail: nadal@lps.ens.fr
and
NESTOR PARGA
Departamento de Física Teórica
Universidad Autónoma de Madrid
Cantoblanco, 28049 Madrid, Spain
E-mail: parga@delta.ft.uam.es
Taking over the original ideas of H. Barlow (1960) and F. Attneave (1954), a systematic approach to the modeling of sensory systems is being developed since the last ten years. The general scheme is as follows:
From step (3), one may also hope to derive plausible learning mechanisms that may lead to this organization, providing models for the epigenetic development.
Step (1) may be relatively easy when dealing with simple animals. For instance one finds motion detectors in the fly visual system, which provide velocity estimations readily used by the motor system. In such case (see e.g. W. Bialek et al, 1991) the Bayesian inference framework (step 2) allows to define the optimal estimator, given the statistics of the signal and the noise level in the receptors (step 3). A different situation is when dealing with, say, the human visual system. Even though independent channels exists, it is clear that many different tasks have to be solved from the same incoming optical flow. One may thus assume that the first layers in the sensory pathway are building a non specific neural representation, or "code", a priori efficient for further processing. Yet, this hypothesis is not enough for specifying an objective cost function (step 2). Indeed, various criteria have been proposed in the literature, among which several based on information theoretic criteria (for an introduction to Information Theory, see e.g. Blahut 1988). The simplest one, studied by various authors, is what has been called the "infomax principle" by R. Linsker (1988): one ask for a neural network which will maximize the mutual information between the output (the neural representation) and the input (say the visual stimuli). The receptors and neural noises, and the finite amount of available resources (number of neurons, synaptic resources) limit the amount of information that can be conveyed by the network on the input, and this limitation renders the maximization a conceptually interesting problem and a generally difficult practical task.
Barlow's proposal was qualitatively different. According to him, it is not only the preservation of information that matters, but more importantly the information presentation: the neural code should be easily readable by the system behind. This imply a compression of information (one should take advantage of the regularities in the stimuli, coding only what makes each stimulus unique), and the search for a code where each neuron is coding for features statistically independent from those coded by the other neurons. These aspects are subsumed in the notion of "redundancy reduction", and the optimal code that achieve redundancy reduction is a factorial code.
It is worth emphazing the differences between these two criteria. Infomax is a quantitative criterion, based on the measure in bits of the statistical relationship between the input and the output of the network. Asking for a factorial code is a qualitative requirement, on the statistical relationship between the output neurons. Departure from factorization is however quantified by a redundancy cost function (to be minimized) expressed in term of information quantities (Barlow 1989, Atick 1992).
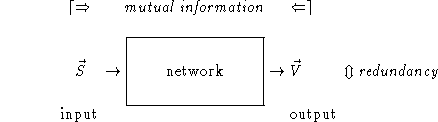
Figure 1: At each time step the environment provides an
input ![]() to the network, which itself produces an output
to the network, which itself produces an output
![]() . The mutual information between the two random
variables
. The mutual information between the two random
variables ![]() and
and ![]() quantifies how much information the output
conveys about the input. The redundancy is a measure of the statistical dependency
between the output activities
quantifies how much information the output
conveys about the input. The redundancy is a measure of the statistical dependency
between the output activities ![]() .
.
The most detailed analytical studies have been performed for simple
feedforward linear
networks, with Gaussian input distributions, for both the infomax
principle (Linsker 1988, van Hateren 1992) and various
implementations of the redundancy reduction
principle (see e.g., Barlow et al 1989, Atick 1992, Redlich 1993, Li and Atick 1994).
Taking account some features specific to particular visual systems,
the results have been applied to the visual system of the fly (van Hateren 1992)
and to mammalian retina (Atick 1992).
Some of these works include a model for the neural code in V1,
which takes into account
altogether contrast, color and motion sensitivities, as well as
stereo-vision, in a multiscale
representation (Li and Atick 1994). The predictions from these calculations are in
qualitative agreement with
known facts on RF of ganglion and V1 cells, and in some cases in
quantitative agreement with
contrast sensitivity curves obtained in psychophysical experiments (Atick 1992). One
can note also that these predicted contrast sensitivity curves are qualitatively very
similar to those obtained from a completly different approach, where the retina is modeled
as a linear electric filter, the architecture of it being based on the detailed knowledge
we have on the retina organization (see J. Herault 1997).
From the theoretical point of
view, what is striking is the extreme similarity between the predictions
derived from these
different criteria: clearly one cannot claim to have pointed out to a basic
organization principle
if other criteria lead to almost identical results! One may think that
the similarity in the
results is due to the use of a linear processing onto a Gaussian
distribution. In fact, one can
see that, in that case of a linear network with Gaussian inputs,
any "reasonable" criterion will lead to a principal component
analysis, with details
depending on the particular constraints under which optimization is
performed. Still, the
linear-Gaussian system remains quite interesting. One can show (Del Giudice et al, 1995)
that the maximization of mutual information, with given input
and output additive
noises, leads to the following features:
However, as soon as one takes into account any nonlinear aspect in the
processing (in particular
the saturation of the transfer functions), one readily finds (Linsker 1988)
that the large freedom we
had in the choice of the solution disappears. It is thus quite important
to understand the role of the nonlinearities.
Furthermore, one may ask for the optimization in the choice of nonlinearities.
On this aspect, a
remarckable work, both theoretical and experimental, has been performed
by Laughlin (1981) on contrast coding in the fly visual system.
Considering a single cell responding to a local contrast (so the input to the
cell is a single scalar), Laughlin derived the optimal transfer function from
an information theoretic point of view, and showed that this prediction
compares very well with
its experimental measurement of the cell respons curve. The theoretical
result is that, in order to maximize the amount of information conveyed
about the input signal, the cell should perform what is known in image processing
as "sampling equalization": for a bounded output activity (say between 0
and 1), the derivative of the transfer function should be equal to the probability
distribution function (p.d.f.) of the input (here the contrast), so that
every possible output occurs with equal probability (Laughlin 1981).
The next step is to consider an array of output neurons, coding for
a multidimensional stimulus. It turns out that a very general statement
can be derived for
feedforward networks with
non linear transfer functions and arbitrary input (signal)
distributions. We have
shown (Nadal and Parga 1994) that, in the low noise limit, the
maximization of mutual information between the input and the output,
if performed over both the synaptic efficacies and the choice of the
transfer functions,
leads to a factorial code - hence to redundancy reduction à la Barlow!
To be more explicit, consider the simplest feedforward network, where each
output neuron has its activity equal to a (specific) nonlinear transfer function
applied to a post-synaptic potential (PSP). This PSP is a linear superposition
of the inputs, the coefficients being the synaptic efficacies. Our result states that
the mutual information will be maximum if: (1) the linear part
of the processing, that is the choice of the synaptic efficacies, is such that
the PSP's (hence the activities of the output cells) are statistically independent; (2) and
for each cell the transfer function is chosen according to the sampling equalization
rule.
Interestingly, this result is related to studies in
signal processing
on blind source separation (BSS) (which is decorrelation in the time
domain, see e.g. Comon, 1994). In particular, it implies
that the mutual information can be used as a cost function
for performing blind source separation (Nadal and Parga 1994). This has been turned into
algorithms showing promising performance on particular BSS applications
(Bell and Sejnowski 1995). It has been then realized that this infomax approach,
for that BSS case, is equivalent to a maximum likelihood approach (Gaeta and Lacoume 1990,
Pham et al 1992),
since they both lead to the very same cost function (Cardoso 1997).
Recently, we have also considered feedforward networks with
arbitrary stochastic output activities (Nadal, Brunel and Parga 1997).
We have shown that the result on factorization
remains valid in the limit of vanishing input noise, whenever it is
the probability distribution
of each output (and not the activity itself) which depends on a (cell dependent)
deterministic function of the input: the maximization of the mutual information
between the input and the output, over the choice of these
deterministic functions, leads to a factorial code.
This is an important extension since it shows that the result
applies in particular in the more realistic case of spiking neurons.
One should emphasis that this factorization, as part of
the optimal solution when
information maximization is performed, occurs also with any non linear
processing before the
output layer. This means that, for any input distribution being a
non linear mixture of
independent spatio-temporal signals, there exists a nonlinear network,
with one or more hidden layers, with which the output will convey as much
information as
possible by providing a factorial representation. This representation,
although not unique,
reflects directly the
statistical structure of the input data. In addition, the infomax cost function,
already tested for the simplest networks
(that is for feedforward networks with no hidden layer, see Bell and Sejnowski 1995), leads
to simple unsupervised backprobagation algorithms
for multilayer networks (Nadal and Parga 1997).
To conclude, we have shown that, at least in the low input noise
limit, maximization of information implies factorization. However,
understanding the possible
consequences of the infomax principle requires a much better
understanding of the statistical
structure of the signal (in the case of the visual system, one may ask
for the structure at the
"pixel" level, or may be at the "object" level.
It remains also to understand what subsists when one consider
noisy inputs. A reasonnable guess is that one will get a result similar
to the one discussed for the linear-Gaussian network: infomax
may lead to redundancy reduction, that is to the factorization of the
input signal into its independent components, together with a redundancy increase
in each one of these independent channels.
Finally, we stress that, although the infomax principle offers a simple
unifying framework for studying sensory coding, it is certainly not the
only possible approach. An interesting alternative approach
has been proposed by Olshausen and Field (1996), who argue that
sparseness might be the relevant quality criterion for the neural code. We note,
however, that the requirement for sparse coding might be considered as a constraint
in an infomax approach, which should be easily taken into account at least
in the linear case
(as we have seen, optimal solutions exists with a large number of ouput cells).
The most recent part of our work presented here is the result of a collaboration
with Nicolas Brunel (LPSENS).
The linear-Gaussian case
It is interesting to see that one may choose a compact solution with
exactly m output neurons,
or distributed solutions with any number (at least equal to m) of
output neurons. This
may be relevant for the understanding of the huge increase in the number
of cells from LGN to V1.
Infomax for a single nonlinear cell
Infomax leads to redundancy reduction
Conclusion
Acknowledgements